Cold-Blooded Monsters or Warm-Hearted Giants?
Predictions of Thermoregulation in Dinosaurs
When we think of reptiles, we think of scaly, cold-blooded creatures, usually relatively inactive between quick darts, often soaking up the sun. When we think of dinosaurs, we tend to think of huge, cold-blooded monsters, more likely feeding or fleeing, hunting or fighting. Dinosaurs were, of course, reptiles themselves; what should strike us about these stereotypic views, however, is perhaps not so much the different life-style we attribute to each, but something even less appreciable than a dinosaur’s habits: the widely shared assumption they were “cold-blooded”. Indeed, the reptiles from which they arose certainly must have been, but dinosaurs gave rise to a fully endothermic lineage – the birds.
So where do dinosaurs fit in? Were they cold-blooded reptiles – like today’s snakes and lizards – or warm-hearted pioneers, like their avian successors?
Since the early seventies, the metabolic rate and thermoregulation of dinosaurs have become topics of hot debate (Bakker, 1968 1), if you’ll pardon the pun. Before then, it was generally assumed that dinosaurs were ectothermic, i.e. their internal body temperature was determined by the environmental conditions at all times. After all, all extant species of reptiles are ectothermic and, powerful as a few may be, need to warm up before they can become fully active. However, it is now irrefutable that birds are direct descendants of dinosaurs (Ostrom, 1973 2; Lloyd et al., 2008 3), and they do possess the metabolic control of their internal body temperature that defines endothermy. So somewhere along the line, dinosaurs would have had to have been “warm-blooded” – whether this feature, however, developed before or after the divergence of avian dinosaurs from the rest of the clade is not so evident.
First of all, it’s important to clarify certain concepts that are key to discussing the thermal physiology of any animal. Although used above, the terms “cold-blooded” and “warm-blooded” are fairly inaccurate and not scientifically rooted: Ectothermic is the correct term to refer to those animals whose body temperature is utterly dependant on the specific environmental conditions at any given time and own behaviour; endothermic animals generate their own internal body heat through a higher metabolic rate and increased aerobic activity. That is not to say that endotherms are not subject to their environment, nor does it imply that ectotherms cannot maintain a constant body temperature; as a matter of fact, the body temperature of endotherms is not necessarily uniform.
Expanding on these grounds, you’ll find that an ectotherm can have a very consistent body temperature if, say, atmospheric temperature is very stable or the animal is very effective at adjusting its behaviour to maintain it (moving between areas of sunlight and shade, for example). Another case in which ectotherm could sustain its internal body temperature is that of inertial homeothermy when, due to a considerable body mass in addition to reasonably stable climatic conditions, the animal doesn’t loose much heat at all. Some endotherms on the other hand, which are generally credited with having strictly stable internal body temperatures, can show great thermal and metabolic fluctuations throughout, for instance, a year – the rises and falls throughout different seasons in, for example, hibernating mammals.
Therefore, there is vast range of potential forms and combinations of thermoregulation mechanisms – and so many different types of endo- and ectothermy. Certainly, there is no simple warm- or cold-blooded division; we could, after all, refer to an inertial homeotherm as being warm-blooded, because it maintains a high body temperature, but they do not generate this heat themselves and are therefore ectothermic at the same time. Given this array of possibilities, why should we preoccupy ourselves trying to prove that dinosaurs were either strictly ectothermic or endothermic, could they not have displayed unique themoregulatory patterns of their own? Many palaeontologists view this as a possibility and indeed as the most likely case (Seebacher, 2003 4).
Endothermy derived from ectothermy, and not directly as a means of thermoregulation as such – in other words, not in response to a need to simply “keep warm” or cool down; it has resulted from selective pressures favouring new adaptations that allowed, for example and amongst other things, greater independence from certain environmental factors, while at the same time increasing their potential level of activity (Hayes and Garland, 1995 5). This does not necessarily imply superiority of endothermy over ectothermy, both forms of thermoregulation have advantages and disadvantages that rely on numerous factors, which is why it is so hard to assess hypothetically whether it would have proven advantageous for dinosaurs to be endothermic, let alone determine whether they actually were or not!
In order to plausibly predict the occurrence of the possible thermoregulatory patterns amongst dinosaurs, the advantages and disadvantages of both endothermy and ectothermy must be put into context and weighed up taking environmental and behavioural factors into consideration, as well as the different shapes, sizes, needs, and niches occupied by the immense diversity of dinosaurian forms throughout their long reign. Of course, although based on information from the fossil record, in dealing with an extinct group of animals it is inevitable that some factors can only be taken into account as assumptions, simplifications, and approximations (Seebacher, 2003 4), but drawing comparisons with living species can prove very useful.
Dinosaurs were a clade of giants. While many moderately sized, small, and even tiny species have been discovered, relatively speaking, dinosaurs were enormous animals as a whole. Being so big holds a range of different implications for an animal’s metabolism. For a start, you need to get big.
Growth rates within all vertebrate groups increase with the body mass of the animals in question (Erickson et al., 2001 6) – a whale, for example, will attain a greater percentage of its final body mass in less time than a mouse will, although the whale will take longer to reach maturity. While this pattern holds true amongst reptiles as well, extant species of this group grow at a much slower rate than birds or mammals. Amongst living vertebrates, birds undoubtedly hold the crown for fastest growth rate (Tjørve, K. M. C. and Tjørve, E., 2010 7).
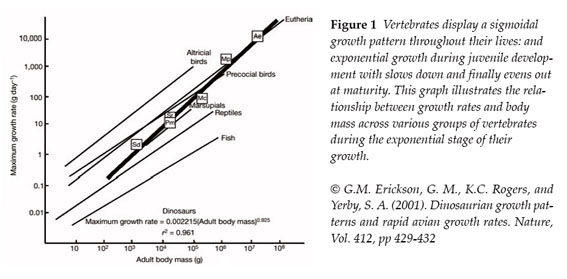
Once again emphasising dinosaur’s huge size, it’s important to note that a higher growth rate than that found in modern reptiles would be necessary in order to achieve it, or dinosaurs would have had to lived to a very ripe old age indeed. Although the concept of longevity should not be shunned on such superficial grounds, there is now conclusive evidence to demonstrate that indeed dinosaurs displayed a growth rate much greater than that of reptiles. This evidence comes from careful histological studies of the bones of various dinosaurs of differing estimated body mass and that of different extant vertebrates. Results from such analysis show that, like other vertebrates, dinosaur growth rate increases with body mass but it is clearly greater that than observed in reptiles alive today (Erickson et al., 2001 6).
How does this relate to thermoregulation? Many authors have considered this fast growth rate as an indication of endothermy, arguing it is more similar to that of endothermic mammals than that of ectothermic reptiles (Benton, 2009 8). However, perhaps surprisingly, the histological studies have also shown that dinosaur growth rate doesn’t even represent an intermediate between reptiles and mammals: i.e. the relation between growth rate and body mass is not merely a faster version than that of living reptiles, nor is is a reduced version of that seen in mammals and birds, but that it displays a pattern of its own [Figure 1] (Erickson et al., 2001 6). Therefore, assuming all dinosaurs were endotherms simply based on a greater similarity between their growth rate and that of extant endotherms rather than other ectotherms, is a little hasty. So these histologic analyses and growth rate studies don’t provide conclusive data on their own.
Another metabolic implication of being big that can potentially effect the thermoregulation of the animal is perhaps a fairly obvious one: it will need a lot of energy (heat) to function properly. In the case of dinosaurs, their likely ecological hierarchies and various clues in the fossil, such as biomechanic studies based on trackways (Alexander, 1976 9), indicate that they composed a very active group, in contrast to extant reptiles which are generally rather sluggish between short bursts of activity. The idea of dinosaurs having the ability to maintain relatively high levels of activity and be able to move quickly without completely exhausting their energetic “allowance”, is one that various authors (Bakker, 1972 1) have used to defend endothermic regulation. After all, it wouldn’t be practical or beneficial if such a large animal as, say, a Tyrannosaurus rex had to wait for the sun to warm up its huge body mass after every hunt – it would be easy to imagine how not very much time for hunting would be left.
However, as mentioned before, environmental factors must also be taken into account. Dinosaurs lived during the Mesozoic, an era when the Earth’s temperature (despite various fluctuations throughout time and geographically) was altogether warmer and the climate milder than it it today. This in mind, and again emphasising the massive size of many species, it is physically possible that a large ectothermic animal could maintain a very stable high body temperature and regulate it simply by moving in and out of sun-exposed and shaded areas, as shown by biophysical studies taking such factors into account (Seebacher, 2003 4). In fact, it is far more likely that large animals regulated their body temperature in this way, as endothermy could have posed more problems than solutions: Elephants today are the largest terrestrial animals, so although they might only equate to what on average would be a medium-sized dinosaur, we can look at their mechanisms of thermoregulation to provide some idea of the advantages and disadvantages of endothermy in large animals.
Elephants live in a hot climate and, as all mammals alive today, they are endothermic, so keeping a high body temperature is not a problem – but avoiding over-heating can be. As a product of millions of years of natural selection, elephants are fully equipped to counteract such effects of environmental heat (their large ears allowing heat loss are one example), but if modern endotherms in warm climates have had to resort to cooling adaptations, it is hard to see how endothermy would be selected for coming from an already effective form of thermoregulation which did not pose such overheating risks in the first place. The selection for endothermic characters in such animals becomes even harder to conceive if we consider an unarguable implication of endothermic regulation: it’s expensive. This extra fee on top of the animal’s energy budget is obviously greater in larger animals than smaller ones, quite simply because they have a greater mass throughout which they must maintain a high metabolic rate. This adds the stress of having to find and process a lot more food. So although it may not have caused an entirely detrimental effect on some animals, if certain traits don’t result in any added benefit, selection pressures will hardly favour them over less costly mechanisms.
At least following these predictions, the extra expenditure of energy isn’t compensated by the benefits of endothermy in animals that are large enough to maintain a high body temperature thanks to their environment and certain behavioural patterns (Seebacher et al., 1999 10). But that’s big animals. Though they represent a smaller portion of the diversity of these reptiles, numerous small dinosaur species thrived throughout the Mesozoic too and, perhaps more importantly, all dinosaurs started off small as babies.
While the ability to maintain a stable internal body temperature may not seem like as much of a priority in smaller dinosaurs, since today’s reptiles get on just fine with their ectothermic ways, it is nonetheless worth studying whether endothermy could have provided any additional benefits which were not applicable to larger specimens, since – as stated above – endothermy must have arisen at some point amongst dinosaurs, it is just unknown whether before or after the differentiation of avian forms.
Being small implies a greater surface to body mass ratio, which means there is a greater area from which to loose the already smaller amount of heat the body possesses, therefore, endothermy could have posed an advantage without the danger of over-heating that large animals would face; smaller endothermic dinosaurs would still have to increase their energy uptake significantly. The extra energy expenditure would again neutralise the potential benefits of endothermy for animals living in a warm climate anyway – they’d probably to fair just fine by obtaining heat from their environment, since their size would mean that although they could loose heat quite rapidly, they could also warm up relatively quickly. So although endothermy wouldn’t necessarily pose any disadvantage for small dinosaurs, current data doesn’t indicate any noticeable advantage either, and therefore natural selection would have no reason to favour traits of an endothermic physiology.
The case may be different, however, for species that lived in colder climates. Although the global climate during Mesozoic was warmer and milder overall than today’s, there was some geographic variation and there it is generally accepted that, at least during some periods, there were seasonal ice caps at the poles (Price, 1999 11). Furthermore, the fossil record shows both the occurrence in high latitudes of large migratory dinosaurs that travelled between these regions and peripheral ones throughout the year and of smaller species that remained there all year round; taking into account the points so far exposed, it is plausible that, if endothermy were to be selected for at all in non-avian dinosaurs, this is the case in which it might prove most advantageous (Seebacher et al., 2003 4).
There is another important aspect to consider when trying to establish whether the cost of producing internal body heat is really what it says on the price-tag: how effectively the heat produced can be conserved. Insulation is key here. The effect of insulation in conserving body heat it enormous. Fossilised impressions of dinosaur skin seem to imply a near scaly texture in most of the samples studied, which are actually relatively few. Currently there is no knowledge of nor data to support any form of dermal insulation in most dinosaurs, except, of course, those of the division that later gave rise to birds.
There are impressions of feathers and feather-like structures in the fossil record that appear long before the appearance of birds, and certainly long before these dinosaurs ever took to the wing (Xu, 2001 12) – if feathers didn’t originally evolve as structures to aid flight, what were they for? One suggestion is certainly as insulation, which makes endothermic regulation seem very plausible amongst dinosaurs that possessed such structures.
Although many more arguments have been proposed in favour and against both endothermy and ectothermy as forms of thermoregulation in dinosaurs (Bakker, 1972 1), weighing up the information that is currently available on this extinct clade of animals with the data obtained about their relevant palaeoenvironments, it is most likely that the group – which included an immense variety of forms and adaptations, sprawled across the globe and, indeed, time – as a whole was neither endothermic nor ectothermic (Seebacher, 2003 4), but that different types of dinosaur possessed a gradient of different types of thermoregulation, from simple ectotherms, through inertial homeotherms and other mid-way stages, to fully endothermic species. It has even been suggested that some of the largest dinosaurs may have displayed a variable thermophysiology in the span of their life-time, switching from an endothermic youth to an inertial homeothermy in adulthood (Benton, 2009 8). Currently, there is avid research being done in the field of the ventilation mechanisms of vertebrates, with particular interest placed on determining the presence or absence of structures known as nasal turbinates in dinosaurs, and the correlation of these respiratory structures with endothermy (Hillenius and Ruben, 2004 13).
For now, however, the strongest evidence seems to support a great diversity of thermoregulation mechanisms amongst dinosaurs, which indeed seems appropriate for such a hugely diverse group of animals, covering everything from cold-blooded monsters to warm-hearted giants.
-------------------------------------
References
1 - Bakker, R. T. (1972). Anatomical and ecological evidence of endothermy in dinosaurs. Nature, 238, 81-85
2 - Ostom, J. H (1973). Ancestry of Birds. Nature, 242
3 - Lloyd, G. T., Davis, K. E., Pisani, D., Tarver, J. E., Ruta, M., Sakamoto, M., Hone, D. W. E., Jennings, R., and Benton, M. J. (2008). Dinosaurs and the Cretaceous Terrestrial Revolution. Proceedings of the Royal Society, Series B 275, 2483-2490.
4 - Seebacher, F. (2003). Dinosaur body temperatures: the occurrence of endothermy and ectothermy. Paleobiology, 29, 105-122
5 - Hayes, J. P., and T. Garland Jr. (1995). The evolution of endothermy: testing the aerobic capacity model. Evolution, 49, 836-847.
6 - G.M. Erickson, G. M., K.C. Rogers, and Yerby, S. A. (2001). Dinosaurian growth patterns and rapid avian growth rates. Nature, 412, 429-432
7 - Tjørve, K. M. C., and Tjørve, E. (2010). Shapes and functions of bird-growth models: how to characterise chick postnatal growth. Zoology, 113 (6), 326-333
8 - Benton, M. J. (2009). Dinosaurs. Current Biology, 9 (8), 318-323
9 - Alexander, R. Mcn. (1976). Estimates of speeds of dinosaurs. Nature, 261, 129-130.
10 - Seebacher, F., G. C. Grigg, and L. A. Beard (1999). Crocodiles as dinosaurs: behavioural thermoregulation in very large ectotherms leads to high and stable body temperatures. Journal of Experimental Biology, 102, 77-86
11 - Price, G. D. (1999). The evidence and implications of polar ice during the Mesozoic. Earth-Science Reviews, 48 (3), 183-210
12 - Xu, X. , Zhou, Z.-H, and Prum, R. O. (2001). Branched integumental structures in Sinornithosaurusand the origin of feathers. Nature, 410, 200–204
13 - Hillenius, W. J., Ruben, J. A. (1999). The Evolution of Endothermy in Terrestrial Vertebrates: Who? When? Why?. Physiological and Biochemical Zoology, 77, 1019-1042
S.-Xaali O'Reilly Berkeley, December 2010
When we think of reptiles, we think of scaly, cold-blooded creatures, usually relatively inactive between quick darts, often soaking up the sun. When we think of dinosaurs, we tend to think of huge, cold-blooded monsters, more likely feeding or fleeing, hunting or fighting. Dinosaurs were, of course, reptiles themselves; what should strike us about these stereotypic views, however, is perhaps not so much the different life-style we attribute to each, but something even less appreciable than a dinosaur’s habits: the widely shared assumption they were “cold-blooded”. Indeed, the reptiles from which they arose certainly must have been, but dinosaurs gave rise to a fully endothermic lineage – the birds.
So where do dinosaurs fit in? Were they cold-blooded reptiles – like today’s snakes and lizards – or warm-hearted pioneers, like their avian successors?
Since the early seventies, the metabolic rate and thermoregulation of dinosaurs have become topics of hot debate (Bakker, 1968 1), if you’ll pardon the pun. Before then, it was generally assumed that dinosaurs were ectothermic, i.e. their internal body temperature was determined by the environmental conditions at all times. After all, all extant species of reptiles are ectothermic and, powerful as a few may be, need to warm up before they can become fully active. However, it is now irrefutable that birds are direct descendants of dinosaurs (Ostrom, 1973 2; Lloyd et al., 2008 3), and they do possess the metabolic control of their internal body temperature that defines endothermy. So somewhere along the line, dinosaurs would have had to have been “warm-blooded” – whether this feature, however, developed before or after the divergence of avian dinosaurs from the rest of the clade is not so evident.
First of all, it’s important to clarify certain concepts that are key to discussing the thermal physiology of any animal. Although used above, the terms “cold-blooded” and “warm-blooded” are fairly inaccurate and not scientifically rooted: Ectothermic is the correct term to refer to those animals whose body temperature is utterly dependant on the specific environmental conditions at any given time and own behaviour; endothermic animals generate their own internal body heat through a higher metabolic rate and increased aerobic activity. That is not to say that endotherms are not subject to their environment, nor does it imply that ectotherms cannot maintain a constant body temperature; as a matter of fact, the body temperature of endotherms is not necessarily uniform.
Expanding on these grounds, you’ll find that an ectotherm can have a very consistent body temperature if, say, atmospheric temperature is very stable or the animal is very effective at adjusting its behaviour to maintain it (moving between areas of sunlight and shade, for example). Another case in which ectotherm could sustain its internal body temperature is that of inertial homeothermy when, due to a considerable body mass in addition to reasonably stable climatic conditions, the animal doesn’t loose much heat at all. Some endotherms on the other hand, which are generally credited with having strictly stable internal body temperatures, can show great thermal and metabolic fluctuations throughout, for instance, a year – the rises and falls throughout different seasons in, for example, hibernating mammals.
Therefore, there is vast range of potential forms and combinations of thermoregulation mechanisms – and so many different types of endo- and ectothermy. Certainly, there is no simple warm- or cold-blooded division; we could, after all, refer to an inertial homeotherm as being warm-blooded, because it maintains a high body temperature, but they do not generate this heat themselves and are therefore ectothermic at the same time. Given this array of possibilities, why should we preoccupy ourselves trying to prove that dinosaurs were either strictly ectothermic or endothermic, could they not have displayed unique themoregulatory patterns of their own? Many palaeontologists view this as a possibility and indeed as the most likely case (Seebacher, 2003 4).
Endothermy derived from ectothermy, and not directly as a means of thermoregulation as such – in other words, not in response to a need to simply “keep warm” or cool down; it has resulted from selective pressures favouring new adaptations that allowed, for example and amongst other things, greater independence from certain environmental factors, while at the same time increasing their potential level of activity (Hayes and Garland, 1995 5). This does not necessarily imply superiority of endothermy over ectothermy, both forms of thermoregulation have advantages and disadvantages that rely on numerous factors, which is why it is so hard to assess hypothetically whether it would have proven advantageous for dinosaurs to be endothermic, let alone determine whether they actually were or not!
In order to plausibly predict the occurrence of the possible thermoregulatory patterns amongst dinosaurs, the advantages and disadvantages of both endothermy and ectothermy must be put into context and weighed up taking environmental and behavioural factors into consideration, as well as the different shapes, sizes, needs, and niches occupied by the immense diversity of dinosaurian forms throughout their long reign. Of course, although based on information from the fossil record, in dealing with an extinct group of animals it is inevitable that some factors can only be taken into account as assumptions, simplifications, and approximations (Seebacher, 2003 4), but drawing comparisons with living species can prove very useful.
Dinosaurs were a clade of giants. While many moderately sized, small, and even tiny species have been discovered, relatively speaking, dinosaurs were enormous animals as a whole. Being so big holds a range of different implications for an animal’s metabolism. For a start, you need to get big.
Growth rates within all vertebrate groups increase with the body mass of the animals in question (Erickson et al., 2001 6) – a whale, for example, will attain a greater percentage of its final body mass in less time than a mouse will, although the whale will take longer to reach maturity. While this pattern holds true amongst reptiles as well, extant species of this group grow at a much slower rate than birds or mammals. Amongst living vertebrates, birds undoubtedly hold the crown for fastest growth rate (Tjørve, K. M. C. and Tjørve, E., 2010 7).
Once again emphasising dinosaur’s huge size, it’s important to note that a higher growth rate than that found in modern reptiles would be necessary in order to achieve it, or dinosaurs would have had to lived to a very ripe old age indeed. Although the concept of longevity should not be shunned on such superficial grounds, there is now conclusive evidence to demonstrate that indeed dinosaurs displayed a growth rate much greater than that of reptiles. This evidence comes from careful histological studies of the bones of various dinosaurs of differing estimated body mass and that of different extant vertebrates. Results from such analysis show that, like other vertebrates, dinosaur growth rate increases with body mass but it is clearly greater that than observed in reptiles alive today (Erickson et al., 2001 6).
How does this relate to thermoregulation? Many authors have considered this fast growth rate as an indication of endothermy, arguing it is more similar to that of endothermic mammals than that of ectothermic reptiles (Benton, 2009 8). However, perhaps surprisingly, the histological studies have also shown that dinosaur growth rate doesn’t even represent an intermediate between reptiles and mammals: i.e. the relation between growth rate and body mass is not merely a faster version than that of living reptiles, nor is is a reduced version of that seen in mammals and birds, but that it displays a pattern of its own [Figure 1] (Erickson et al., 2001 6). Therefore, assuming all dinosaurs were endotherms simply based on a greater similarity between their growth rate and that of extant endotherms rather than other ectotherms, is a little hasty. So these histologic analyses and growth rate studies don’t provide conclusive data on their own.
Another metabolic implication of being big that can potentially effect the thermoregulation of the animal is perhaps a fairly obvious one: it will need a lot of energy (heat) to function properly. In the case of dinosaurs, their likely ecological hierarchies and various clues in the fossil, such as biomechanic studies based on trackways (Alexander, 1976 9), indicate that they composed a very active group, in contrast to extant reptiles which are generally rather sluggish between short bursts of activity. The idea of dinosaurs having the ability to maintain relatively high levels of activity and be able to move quickly without completely exhausting their energetic “allowance”, is one that various authors (Bakker, 1972 1) have used to defend endothermic regulation. After all, it wouldn’t be practical or beneficial if such a large animal as, say, a Tyrannosaurus rex had to wait for the sun to warm up its huge body mass after every hunt – it would be easy to imagine how not very much time for hunting would be left.
However, as mentioned before, environmental factors must also be taken into account. Dinosaurs lived during the Mesozoic, an era when the Earth’s temperature (despite various fluctuations throughout time and geographically) was altogether warmer and the climate milder than it it today. This in mind, and again emphasising the massive size of many species, it is physically possible that a large ectothermic animal could maintain a very stable high body temperature and regulate it simply by moving in and out of sun-exposed and shaded areas, as shown by biophysical studies taking such factors into account (Seebacher, 2003 4). In fact, it is far more likely that large animals regulated their body temperature in this way, as endothermy could have posed more problems than solutions: Elephants today are the largest terrestrial animals, so although they might only equate to what on average would be a medium-sized dinosaur, we can look at their mechanisms of thermoregulation to provide some idea of the advantages and disadvantages of endothermy in large animals.
Elephants live in a hot climate and, as all mammals alive today, they are endothermic, so keeping a high body temperature is not a problem – but avoiding over-heating can be. As a product of millions of years of natural selection, elephants are fully equipped to counteract such effects of environmental heat (their large ears allowing heat loss are one example), but if modern endotherms in warm climates have had to resort to cooling adaptations, it is hard to see how endothermy would be selected for coming from an already effective form of thermoregulation which did not pose such overheating risks in the first place. The selection for endothermic characters in such animals becomes even harder to conceive if we consider an unarguable implication of endothermic regulation: it’s expensive. This extra fee on top of the animal’s energy budget is obviously greater in larger animals than smaller ones, quite simply because they have a greater mass throughout which they must maintain a high metabolic rate. This adds the stress of having to find and process a lot more food. So although it may not have caused an entirely detrimental effect on some animals, if certain traits don’t result in any added benefit, selection pressures will hardly favour them over less costly mechanisms.
At least following these predictions, the extra expenditure of energy isn’t compensated by the benefits of endothermy in animals that are large enough to maintain a high body temperature thanks to their environment and certain behavioural patterns (Seebacher et al., 1999 10). But that’s big animals. Though they represent a smaller portion of the diversity of these reptiles, numerous small dinosaur species thrived throughout the Mesozoic too and, perhaps more importantly, all dinosaurs started off small as babies.
While the ability to maintain a stable internal body temperature may not seem like as much of a priority in smaller dinosaurs, since today’s reptiles get on just fine with their ectothermic ways, it is nonetheless worth studying whether endothermy could have provided any additional benefits which were not applicable to larger specimens, since – as stated above – endothermy must have arisen at some point amongst dinosaurs, it is just unknown whether before or after the differentiation of avian forms.
Being small implies a greater surface to body mass ratio, which means there is a greater area from which to loose the already smaller amount of heat the body possesses, therefore, endothermy could have posed an advantage without the danger of over-heating that large animals would face; smaller endothermic dinosaurs would still have to increase their energy uptake significantly. The extra energy expenditure would again neutralise the potential benefits of endothermy for animals living in a warm climate anyway – they’d probably to fair just fine by obtaining heat from their environment, since their size would mean that although they could loose heat quite rapidly, they could also warm up relatively quickly. So although endothermy wouldn’t necessarily pose any disadvantage for small dinosaurs, current data doesn’t indicate any noticeable advantage either, and therefore natural selection would have no reason to favour traits of an endothermic physiology.
The case may be different, however, for species that lived in colder climates. Although the global climate during Mesozoic was warmer and milder overall than today’s, there was some geographic variation and there it is generally accepted that, at least during some periods, there were seasonal ice caps at the poles (Price, 1999 11). Furthermore, the fossil record shows both the occurrence in high latitudes of large migratory dinosaurs that travelled between these regions and peripheral ones throughout the year and of smaller species that remained there all year round; taking into account the points so far exposed, it is plausible that, if endothermy were to be selected for at all in non-avian dinosaurs, this is the case in which it might prove most advantageous (Seebacher et al., 2003 4).
There is another important aspect to consider when trying to establish whether the cost of producing internal body heat is really what it says on the price-tag: how effectively the heat produced can be conserved. Insulation is key here. The effect of insulation in conserving body heat it enormous. Fossilised impressions of dinosaur skin seem to imply a near scaly texture in most of the samples studied, which are actually relatively few. Currently there is no knowledge of nor data to support any form of dermal insulation in most dinosaurs, except, of course, those of the division that later gave rise to birds.
There are impressions of feathers and feather-like structures in the fossil record that appear long before the appearance of birds, and certainly long before these dinosaurs ever took to the wing (Xu, 2001 12) – if feathers didn’t originally evolve as structures to aid flight, what were they for? One suggestion is certainly as insulation, which makes endothermic regulation seem very plausible amongst dinosaurs that possessed such structures.
Although many more arguments have been proposed in favour and against both endothermy and ectothermy as forms of thermoregulation in dinosaurs (Bakker, 1972 1), weighing up the information that is currently available on this extinct clade of animals with the data obtained about their relevant palaeoenvironments, it is most likely that the group – which included an immense variety of forms and adaptations, sprawled across the globe and, indeed, time – as a whole was neither endothermic nor ectothermic (Seebacher, 2003 4), but that different types of dinosaur possessed a gradient of different types of thermoregulation, from simple ectotherms, through inertial homeotherms and other mid-way stages, to fully endothermic species. It has even been suggested that some of the largest dinosaurs may have displayed a variable thermophysiology in the span of their life-time, switching from an endothermic youth to an inertial homeothermy in adulthood (Benton, 2009 8). Currently, there is avid research being done in the field of the ventilation mechanisms of vertebrates, with particular interest placed on determining the presence or absence of structures known as nasal turbinates in dinosaurs, and the correlation of these respiratory structures with endothermy (Hillenius and Ruben, 2004 13).
For now, however, the strongest evidence seems to support a great diversity of thermoregulation mechanisms amongst dinosaurs, which indeed seems appropriate for such a hugely diverse group of animals, covering everything from cold-blooded monsters to warm-hearted giants.
-------------------------------------
References
1 - Bakker, R. T. (1972). Anatomical and ecological evidence of endothermy in dinosaurs. Nature, 238, 81-85
2 - Ostom, J. H (1973). Ancestry of Birds. Nature, 242
3 - Lloyd, G. T., Davis, K. E., Pisani, D., Tarver, J. E., Ruta, M., Sakamoto, M., Hone, D. W. E., Jennings, R., and Benton, M. J. (2008). Dinosaurs and the Cretaceous Terrestrial Revolution. Proceedings of the Royal Society, Series B 275, 2483-2490.
4 - Seebacher, F. (2003). Dinosaur body temperatures: the occurrence of endothermy and ectothermy. Paleobiology, 29, 105-122
5 - Hayes, J. P., and T. Garland Jr. (1995). The evolution of endothermy: testing the aerobic capacity model. Evolution, 49, 836-847.
6 - G.M. Erickson, G. M., K.C. Rogers, and Yerby, S. A. (2001). Dinosaurian growth patterns and rapid avian growth rates. Nature, 412, 429-432
7 - Tjørve, K. M. C., and Tjørve, E. (2010). Shapes and functions of bird-growth models: how to characterise chick postnatal growth. Zoology, 113 (6), 326-333
8 - Benton, M. J. (2009). Dinosaurs. Current Biology, 9 (8), 318-323
9 - Alexander, R. Mcn. (1976). Estimates of speeds of dinosaurs. Nature, 261, 129-130.
10 - Seebacher, F., G. C. Grigg, and L. A. Beard (1999). Crocodiles as dinosaurs: behavioural thermoregulation in very large ectotherms leads to high and stable body temperatures. Journal of Experimental Biology, 102, 77-86
11 - Price, G. D. (1999). The evidence and implications of polar ice during the Mesozoic. Earth-Science Reviews, 48 (3), 183-210
12 - Xu, X. , Zhou, Z.-H, and Prum, R. O. (2001). Branched integumental structures in Sinornithosaurusand the origin of feathers. Nature, 410, 200–204
13 - Hillenius, W. J., Ruben, J. A. (1999). The Evolution of Endothermy in Terrestrial Vertebrates: Who? When? Why?. Physiological and Biochemical Zoology, 77, 1019-1042
S.-Xaali O'Reilly Berkeley, December 2010



Comments
Post a Comment